�c������, ���Ǝq, ���앐�N�D�@�o�������b�g���玿�̖��o���������D���{���Ɠ��w�
15: 337-340, 2008. �m���ǗL�n
�ړI
��̖��Q�Ŋ��ꂽ���o���́A�ۍ��_�o�A����_�o�Ȃǂ̖����_�o����Ė��o�̑�ꎟ���p�j�ł��鉄���Ǒ��j�֑�����1)�B���b�g�ł́A���̏��͂���ɁA�������r�T�j�A�����㕠�����j���זE���ւƏ��������ď�������A�ŏI�I�ɑ�]�玿���o�̖�ɓ��B�����Ƃ��Ă̒m�o��������ƍl������1)�B���m�o�̐_�o�@�\�Ɋւ��錤���̑����́A�����_�o�A�����Ǒ��j����ы������r�T�j��ΏۂƂ��Ă���A�����㕠�����j���זE�����]�玿���o�̖��ΏۂƂ������̂͏��Ȃ��B�܂��]���A���o����@�\�̌����ł́A�o�����Ă��铮�����g�p���邱�Ƃ��d�v�ł���Ǝw�E����Ă������A�o�����̃��b�g��p������]�玿���o�̖�Ɋւ���j���[�������x���̌����͂قƂ�ǂȂ��̂�����ł���B�{�����̖ړI�́A���̍ō��ʒ����ł����]�玿���o�̖�ɂ����閡�o���̕\���l�����A�o�������b�g��p�����j���[�������x���̌����Ŗ��炩�ɂ��邱�Ƃł���B
���@
���b�g���y���g�o���r�^�[���������i35
mg/kg�Aip�j�Ŕ]��ʌŒ葕�u�ɌŒ肵�A���W����ɖ��������������ɌŒ�p����A�_�v�^�[�����邽�߂̃z���_�[���r�X�Ǝ��ȗp�Z�����g�ɂ��`������2)�B�܂������ɁA���o�n�t�����p�̌��o���J�e�[�e���̗��u���s�����B��p�����A���b�g�Ɉ����������{���A������]��ʌŒ葕�u�ɌŒ肵����ԂŁA���o���J�e�[�e�����璍����������n�t��ێ悷�邱�Ƃ��P�������B
�o�����ł̋L�^���ɂ́A�P�����Ɠ��l�Ɉ������������ɂ��������b�g�̓��玿�i��]�玿���o�̖�j�ɃK���X�핢�^���O�X�e���d�ɂ��h�����A���X�ɓd�ɂ�[���ւƐi�߂Ȃ���_�o�������L�^�����B�P��j���[���������ɕ����\�ȋL�^������ꂽ��A���̈ʒu�œd�ɂ��~�߁A���o���J�e�[�e�����琅�□�o�n�t�𒍓����A�����t�̐ێ掞�̊����ω��i�����j���L�^�E��͂����B�Ȃ��A���o�n�t�́A�Ö��Ƃ���0.3
M �n�t�A�����Ƃ���0.1
M�H�����A�_���Ƃ���0.001
M�N�G���_�n�t�A����ыꖡ�Ƃ���0.0001M���_�L�j�[�l�n�t��p����3,4)�B
����
�P)�L�^�j���[����������щ����j���[�����̕��ށF�o�������b�g�̓��玿���瑍��45�̃j���[�����������L�^���A���̓�22��(48.9%)�����o������������(���o�����j���[����)�B����疡�o�����j���[�����́A�ł������������閡�o�n�t�̎�ނ��A�H���x�X�g�j���[����(7��)�A�x�X�g�j���[����(6��)�A�N�G���_�x�X�g�j���[����(5��)�A����щ��_�L�j�[�l�x�X�g�j���[����(4��)�ɕ��ނł����B�܂��A45�̒��ɂ͐��ɑI��I�ɉ�������j���[�����□�o�h����̐�ɓ��ٓI�ɉ�������j���[�������������B
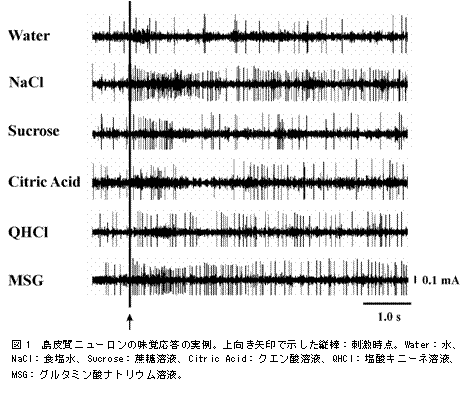
�Q)���o�����j���[�����̎���Ɩ��o�����j���[�����Q�̉����v���t�@�C���F �}1�ɂ́A�H���x�X�g�j���[�����̉�����������Ă���B���̃j���[�����̊����́A���b�g�ɐ���^���Ă��w�ǂ͕ω����Ȃ���(Water)�A�H����(NaCl)��O���^�~���_�i�g���E���n�t(MSG)��^����Ƌ}���ɑ��������B��(Sucrose)�A�N�G���_(Citric
Acid)����щ��_�L�j�[�l(QHCl)�̊e�n�t��^�����ꍇ�́A���^����ɂ킸���Ɋ����������������A�n�t��N�G���_�n�t�ł͂��̌㊈���̗}���������Ԃ��ώ@���ꂽ�B
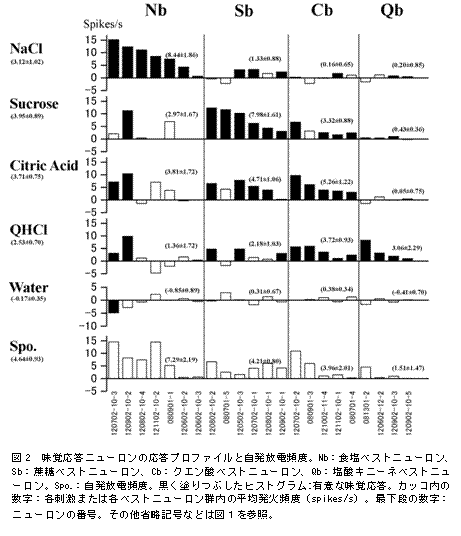
�}2�ɂ́A����L�^����22�̖��o�����j���[�����̊e���o�n�t�ւ̉������x(���o�h����5�b�Ԃ̕��ϕ��d�p�x-�����^��5�b�Ԃ̕��ϕ��d�p�x)�A���ւ̉������x(�����^��5�b�Ԃ̕��ϕ��d�p�x-�����^�O2�b�Ԃ̕��ώ������d�p�x)����ю������d�p�x���q�X�g�O�����Ƃ��Ď����Ă���B���̐}�ł͍����炻�ꂼ��A�H���A�A�N�G���_�A���_�L�j�[�l�̊e�x�X�g�j���[�����Q���Ƃɕ��ׂĂ���A�܂������e�x�X�g�j���[�����Q���ł͉������̋������̂قǍ��ɗ���悤�ɕ��ׂĂ���B�S�̓I�ȌX���Ƃ��ĐH���x�X�g�j���[�����Ɖ��_�L�j�[�l�x�X�g�j���[�����ł͍œK���o�h��(���ꂼ��A�H�����Ɖ��_�L�j�[�l�n�t)�ȊO�̖��o�h���ɂ͂��܂艞�����Ȃ����A�x�X�g�j���[�����ł��n�t�ւ̉����ƃN�G���_�n�t�ւ̉�������r�I���Ă���A�܂��A�N�G���_�x�X�g�j���[�����ł́A�H�����ȊO�̗n�t(�A�N�G���_�A���_�L�j�[�l�̊e�n�t)�ʼn�������r�I�悭���Ă����B
�l�@
�{�����ł́A�o���E�s�������b�g�̓��玿�ɂ����閡�o���̕\���l������͂����B�����Ǒ��j�A�������r�T�j�A�����㕠�����j���זE���Ȃǂ̖��o�̈�ł́A��r�I�[���l���u�^�[���������ł����o�U���������L�^�ł���̂ɑ��āA���玿�ł̓l���u�^�[���������ł͖��ĂȖ��o�U���������قƂ�NjL�^�ł��Ȃ��B���̂��߁A���玿�ɂ����閡�o�U�������̋L�^�ł́A�]���A�E���^��5-7)�-�N�������[�X8)�Ȃlj����}���̂��܂苭���Ȃ��Ƃ���閃���g�p����Ă����B�����������̖�����ł��A���������[�x���[���Ȃ�Ɩ��o�U�������͖w�NjL�^�ł��Ȃ��Ȃ�i�����\�f�[�^�j�B����ɑ��Ė{�����ŗp�����o�������b�g�ł́A�������ɂ͋L�^�ł��Ȃ������]��ʍ��W�ł͂�����Ƃ������o�U����������і��o�j���[�����������L�^���邱�Ƃ��ł����B�]���āA���玿�ɂ����閡�o���̕\���l���Ɋւ��錤���ł́A�o�����̓�����p���邱�Ƃ̗��_�����ɑ傫���Ƃ�����B����������́A�K�������������̌�����ے肷����̂ł͂Ȃ��B�Ⴆ�A�o�����ł́A�@���o���ւ̖��o�n�t���^�̃R���g���[��������ł���A�A�n�t��ێ悷�邽�ߖ����ȂǗn�t�ێ�ɂ��e���������ł��Ȃ��A�B������(2���Ԉȏ�)�ɂ킽���ē������S�����邱�Ƃ͍���ł���Ȃǂ̌��_������B�����ړI�ɏ]���čœK�Ȏ����n��p���A�������Ɗo���s�����̎����̃����b�g�������������s�����Ƃ��d�v�ł��낤�B
���玿�j���[�����̖��o�������x�́A�����Ǒ��j3)�⋴�����r�T�j4)�̃j���[�����Ɣ�r���Ďォ�����B�ߋ��̌����ŁA���o�������x�͎����㕠�����j���זE���̃��x���ł��łɉ����⋴�Ɣ�r����ƌ��シ�邱�Ƃ��m���Ă���9)�A����̌��ʂƈ�v���Ă����B���̂��Ƃ́A���̃��o�[�X�g�l�X�����ʂ̖��o���p�j�����ቺ���邱�Ƃ��Ӗ����Ă���A��]�玿�Ŗ��o����Ȃ��Ȃ��悤�ɂ��邽�߂ɂ́A���炩�̑㏞�@�\������Ǝv����B�܂��A���玿�j���[�����̖��o�����h���I�𐫂́A�����Ǒ��j�Ɠ����x�ł��������A�������r�T�j���͒Ⴉ�����B����ɁA���ʒ��p�j�̖��o�����́A�������x�̍��͂����Ă��A������̖��o�n�t�ɑ��Ă��h���Ɠ����Ɋ������㏸�����ԂƂƂ��Ɋ����ቺ����Ƃ�����r�I�ꗥ�Ȏ��Ԍo�߂̃p�^�[�������X��������5,6)�B����ɑ��āA���玿�j���[�����́A�}1�ɂ��������悤�ɁA���o�h���̎�ނɂ�芈���̌o���ω��i����/�}���p�^�[���j���قȂ�X��������A�]���ē��玿�ł͖��o����ϔ��Η�����łȂ��X�p�C�N�^�C�~���O�Ƃ��Ă�����������Ă���\��������B�����̎����́A���o������K�w�I�ɐi�݁A�����̈�قǂ�蕡�G�Ȗ��o�������\������Ă��邱�Ƃf���Ă���̂�������Ȃ��B
����
1) Lundy RF and Norgren
R: Gustatory system. In The Rat Nervous System. Paxinos
G (ed), Elsevier Academic Press,
2) Norgren R, Nishijo H and Travers SP: Taste responses from the entire
gustatory apparatus.
3) Nakamura K and Norgren
R: Gustatory responses of neurons in the nucleus of the solitary tract of
behaving rats. J. Neurophysiol. 66: 1232-1248
(1991)
4) Nishijo H
and Norgren R: Responses from parabrachial
gustatory neurons in behaving rats. J. Neurophysiol.
63: 707-724 (1990)
5) Yamamoto T, Yuyama
N, Kato T and Kawamura Y: Gustatory responses of cortical neurons in rats. I.
Response characteristics. J. Neurophysiol. 51:
616-635 (1984)
6) Ogawa H, Ito S, Murayama
N and Hasegawa K: Taste area in granular and dysgranular
insular cortices in the rat identified by stimulation of the entire oral
cavity. Neurosci. Res. 9: 196-201 (1990)
7) Kosar E,
Grill HJ and Norgren R: Gustatory cortex in the rat.
I. Physiological properties and cytoarchitecture. Brain
Res. 379: 329-341 (1986)
8) Cechetto DF
and Saper CB: Evidence for a viscerotopic
sensory representation in the cortex and thalamus in the rat. J. Comp. Neurol. 262: 27-45 (1987)
9) Norgren R:
Central neural mechanisms of taste. In Handbook of Physiology - The
nervous system III. Sensory Processes. Brookhart JM, Mountcastle VB, Darian-Smith I,
and Geiger SR (eds), American Physiological Society
Bethesda, pp.1087-1128 (1987)