- A. 海馬体と記憶
-
図1はヒトの脳の模式図ですが,ヒトも含め霊長類では海馬体は側頭葉の内側部に位置し,記憶形成と一定期間の保持に重要な脳領域です.海馬体を含む側頭葉内側部に損傷のある患者さんでは健忘症が起こりますが,受傷後に新たな記憶を形成できなくなる前行健忘と,受傷以前約2年に及ぶ逆行健忘の両方が見られます.また逆行健忘に関しては,受傷時点に近いほど重篤であるという時間勾配があるのも特徴です.サルで海馬体を破壊すると,ヒトの健忘症に類似した症状,すなわち前行健忘と約2ヶ月間(人の場合と比較するとかなり短い)に及ぶ,やはり時間勾配のある逆行健忘が起こります.こうした事実から海馬体は記憶の首座とされてきましたが,海馬体が損傷しても障害の起こる記憶と起こらない記憶があることも知られています.海馬体損傷により,いつ,どこで,だれが(あるいは何が),どうしたといった出来事の記憶(エピソード記憶)は障害されますが,技能や習慣など,いわゆる“からだで覚える”ことは正常人と同じようにできます.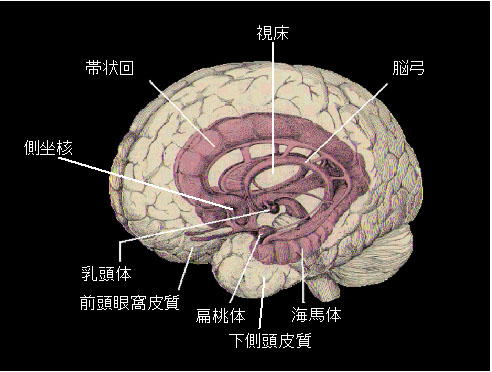
図1 ヒトの脳の模式図. - B. 刺激位置識別応答ニューロン
-
さて,私が実際に行ってきた研究に話を進めていくことにします.先に述べましたように,海馬体依存的であるエピソード記憶は,「いつ,どこで」という時間的・空間的に定位される記憶ですが,わたくしはこのうち,まず空間という切り口から,海馬体による情報処理機構解明にアプローチしました.図2は,空間情報処理に関る海馬体ニューロンの応答例で,私が大学院生として実験を始めてまもなく見つけたものです.この実験では,サルの海馬体に微小電極を刺入して個々のニューロンの電気的活動(細胞外の活動電位,すなわちインパルス放電)を記録した状態で,様々な視覚刺激や聴覚刺激を異なる方向からサルに呈示し,活動の変化の仕方(応答様式)を解析しました.たとえば,視覚刺激としてリンゴ,注射器,小さな棒などを,左前方,前方,右前方から見せたり,聴覚刺激として1,000Hzの純音を左前方,前方,右前方,左後方,後方,右後方の6方向から聞かせたりしました.このニューロンは,右前方から視覚刺激が呈示されると,呈示物体の種類とは無関係に活動増加(促進応答)しました.同じ物体でも他の方向から呈示したときには応答せず,また右前からでも聴覚刺激に対しては有意の応答はありません.従いまして,このニューロンは,呈示物体の位置を視覚的にコードしているわけです(刺激位置識別応答ニューロン)1).このような刺激位置識別応答ニューロンの性質をもう少し詳しく調べるとさらに面白いことがわかってきました.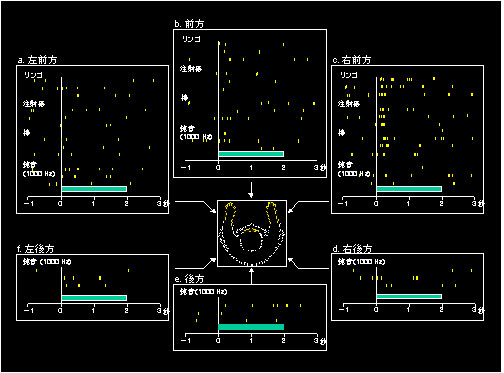
図2 刺激位置識別応答ニューロンの応答例.
ラスター表示:各々の小さな縦線はニューロンのインパルス放電(活動電位).横軸:時間,0の時点:刺激呈示の開始.横軸上の横棒:刺激呈示期間.
図3Aも刺激位置識別応答ニューロンの例ですが,実験室の中でのサルの向きがニューロン応答にどのような影響を与えるかを調べました.実験室ではサルは通常図3Aaに示してある方向を向いていますが,その場合このニューロンは,右前からの刺激呈示に対して強く応答しています.サルの向きを左回り(Ab)または右回り(Ac)に45度回転しても,サル自身から見て常に右の方から呈示された刺激に対してだけ応答しました.従ってこのニューロンはサル自身の頭を中心とした(自己中心)座標系で呈示刺激の位置をコードしていることがわかりました.図3Bはまた別の刺激位置識別応答ニューロンの例ですが,サルの向きとは無関係に,部屋の右前の場所から物体を呈示すると応答があります.従ってこのニューロンは,呈示刺激の位置を外界空間の座標系,いいかえれば,実験室という基準フレームででコードしていることがわかります.私たちは,この研究から海馬体における空間情報処理として,自己中心的座標系や外界空間座標系により呈示刺激の位置をコードしていることを明らかにしました1, 2).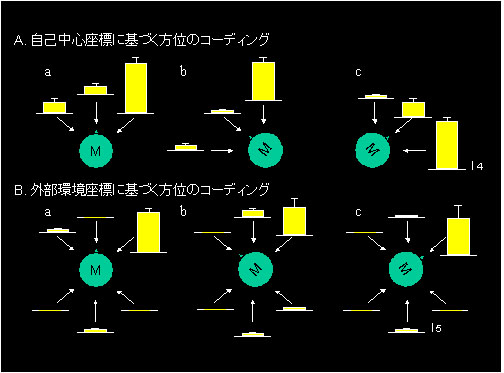
図3 自己中心座標系(A)または外界環境座標系(B)に基づき刺激位置をコードするニューロンの応答例.
各棒グラフと誤差線:3回の刺激呈示に対する平均応答強度と平均誤差.M:サル. - C. 場所ニューロン
-
サルの海馬体にはサル自身の居場所をコードするニューロン(場所ニューロン)も存在します3).図4Aには,この研究で用いた実験装置の模式図を示してありますが,一種のサル用の自動車です.サルはフロントパネルに設置してあるレバーを操作することにより2.5m x 2.5mの範囲を移動できます.図4Bはサルが“自動車”を運転した移動軌跡であり,番号の順に移動しました.図4Cは,場所ニューロンの例で,サルが右前(図4B,場所1-3)にいるときに強く活動していることがお分かりになると思います.また私たちは,サルの移動様式が場所ニューロン活動に影響することも明らかにしました4).図5には,図4と同じニューロンを,移動様式を変えて応答性がどうなるかを調べた結果を示してあります.図5AとBは,サルが自分自身でレバーを操作し,反時計回り(A)または時計回り(B)に自動車を運転したとき,図5Cはサル自身ではなく,実験者がコンピュータ制御によりサルを強制的に反時計回りに移動させたときのニューロン活動です.サル自身で移動したときには図6と同じように右前で強く活動していますが,強制的に移動させられたときにはその活動が見られません.すなわち海馬体ニューロンが場所特異的に応答するためには,動物が自らの意思で移動することが重要な条件となります.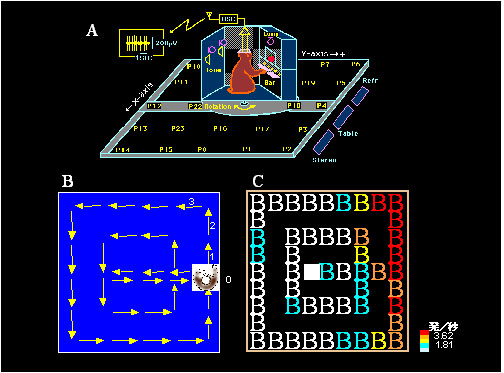
図4 サル海馬体場所ニューロンの応答例.
A:実験装置(“サルの自動車”)の模式図.前面パネルには5個のレバー(Bar)が設置してあり,サルがレバーを押すと自動車が前後左右に移動.微小電極から記録されたニューロン活動はテレメータ(osc)により送信される.P0 – 24:自動車の移動範囲を5 x 5(25)分割したときの各々の場所.
B:サルの移動経路の一例.矢印:移動方向.場所0(上記AにおけるP0)から出発.
C:ニューロン活動(各場所における平均放電頻度)のカラーマップ.このニューロンは,場所1-3で活動を増加した. -

図5 海馬体場所ニューロンの移動様式依存性.
AとB:サルが自分自身でレバーを操作し,反時計回り(A, ccw)または時計回り(B, cw)に自動車を運転.
C:サル自身ではなく,実験者がコンピュータ制御によりサルを強制的に反時計回り(ccw)に移動.
その他の説明は図4を参照. - D. 海馬体脳波と動物の移動様式
-
上述の「自らの意思で移動することが海馬体の活動に重要である」との知見は,最近私たちが行った別の種類の実験からも支持されています.図6は典型的な海馬体脳波を示してありますが,ラットなどのげっ歯類では,探索行動下,ウレタンなどの麻酔下,レム睡眠中などには周期性徐波活動(または海馬体シータ波)と呼ばれる高振幅で規則正しい6~10Hzの正弦波様の脳波が顕著に出現します(図6A).一方ラットがじっとしている時,拘束されている時,摂食・飲水時,ノンレム睡眠時などには,高振幅不規則波と呼ばれ脳波が出現します(図6B).周期性徐波と動物の移動様式との間には,“ラット自らが自由に歩行しているときには周期性徐波が現われるが,拘束された状態で受動的に位置移動させられても出現しない”という関係があることが知られています.しかし,ヒトを含め霊長類の海馬体は脳の深部にあり,電極を正確に刺入することが極めて困難であるため,これまで海馬体脳波に関する研究は少なく,とくに動物の移動様式との関係についてはまったく調べられていませんでした.そこで私たちは,磁気共鳴画像法を用いて電極の刺入精度を高めることによりサルの海馬体に正確に電極を刺入し(図7)5),自由に行動しているサルの海馬体から脳波記録を試みました.図8には,その結果を示してあります.サルが歩行を始めると典型的な高振幅の周期性徐波が出現しましたが,じっとしている時,摂食・飲水時,また拘束された状態で受動的に位置移動させられているときには周期性徐波は出現せず,従ってげっ歯類で見られる周期性徐波と類似した性質のあることがわかってきました.しかし,げっ歯類とは異なる点も見つかり,たとえば,①サルの周期性徐波は約3~6Hzで(図8B),これはラットにおける典型的な周期性徐波の周波数帯(8 Hz 前後)と比較して低いこと,②ラットでは歩行により周期性徐波が必発するが,サルでは出現確率は上がるものの歩行時に常に見られるとは限らないこと,③ラットでは特定の行動時に連続的に周期性徐波が現れるのに対し,サルでは一過性に現れその後不明瞭になることなども明らかになっています6).このように,海馬体はサルが能動的に移動しているときに活性化することがわかってきましたので,現在は自由行動下のサルの海馬体からニューロン活動と脳波を同時記録し,周期性徐波の場所情報処理における意義を明らかにしようとしています.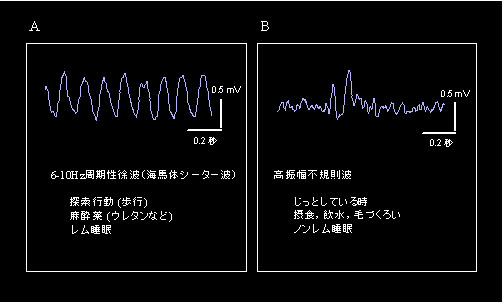
図6 げっ歯類(ラット)の海馬体脳波
A:周期性徐波活動.
B:高振幅不規則波. -
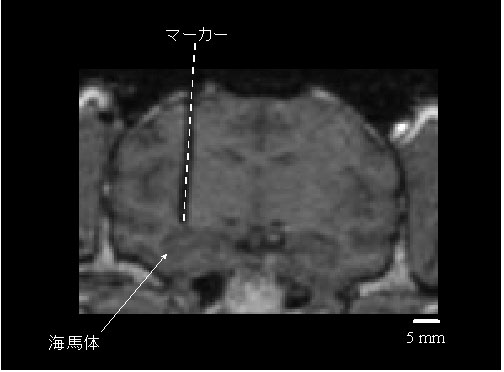
図7 サル脳の海馬体を含む冠状断面の磁気共鳴画像.
タングステンロッド(直径0.5 mm)のマーカー(点線部分)を刺入して撮像.
撮像:Magnetom Vision 1.5 T (Siemens, Germany), 3D FLASH法(TR, 20 ms; TE, 5 ms; flip angle, 30°; FOV, 25.6 cm; matrix 256 x 256; スライス厚,1 mm; スライス間ギャップ,なし).方法の詳細はAsahi et al., 2003を参照.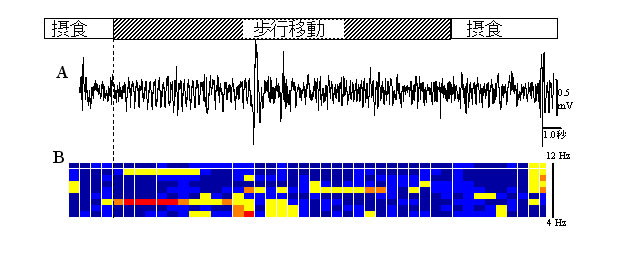
図8 海馬体脳波と動物の移動様式.
A:実際の脳波記録.横軸,時間;縦の点線,歩行開始時点;縦軸,電位振幅.
B:記録脳波を短期高速フーリエ変換(short FFT)処理した結果のカラー表示.横軸,時間(時間スケールはAと同じ);縦軸,周波数.カラー表示:各周波数帯におけるパワー.
|
|
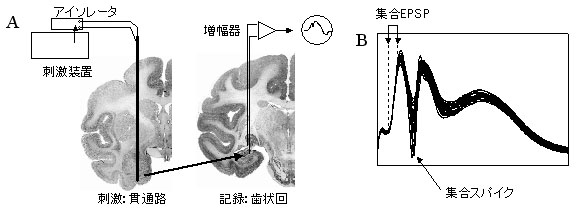
図9 サル海馬体からの誘発電位記録. A:実験の模式図.海馬体の入力線維である貫通路を電気刺激し,歯状回から誘発電位を記録. B:実際の誘発電位記録波形(10波形の重ね合わせ).1対(2本)矢印で示した誘発電位部分:集合興奮性シナプス後電位(EPSP)成分,一本矢印で示した誘発電位部分:集合スパイク成分. |
|
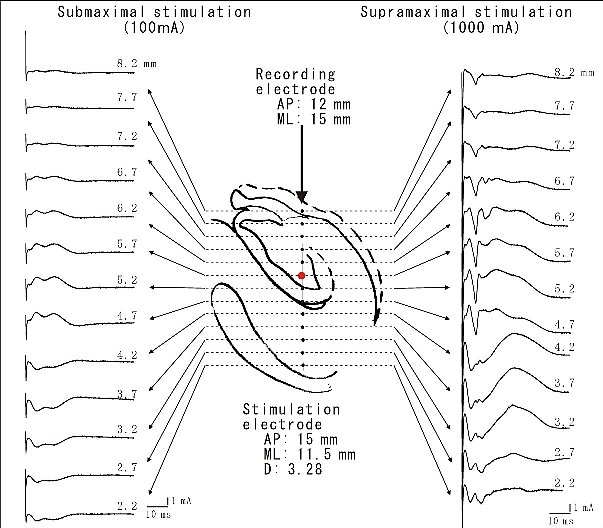
| 図10 |
|
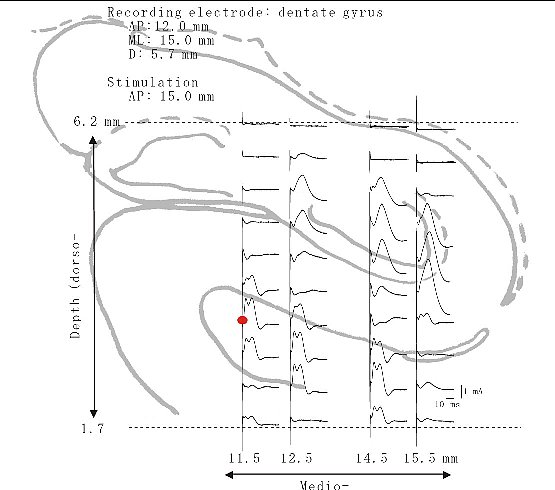
| 図11 |
|
|
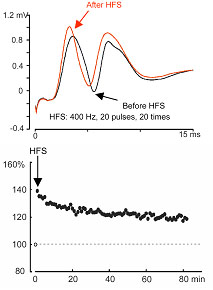
図12 反復刺激後長期増強 A:実際の誘発電位波形.黒線:高頻度刺激前,赤線:高頻度刺激後. B:誘発電位のEPSPスロープの時間経過.白丸:高頻度刺激前の10分間のデータの平均,黒丸:高頻度刺激後の1分ごとのデータ. |